









Servicios
 Como citar este trabajo
Como citar este trabajoAn. 3. Enc. Energ. Meio Rural 2003
Disponibilidade de energia na forma de calor da biomassa lenhosa de um cerrado sensu stricto da região de Brasília
Ailton Teixeira do ValeI; Maria Aparecida Mourão BrasilII; Alcides Lopes LeãoII
IDepartamento de Engenharia Florestal, Faculdade de Tecnologia, Universidade de Brasília,70.910-900, Brasília, DF, tel.: (061) 273 6026, fax: (061) 347 5458
IIDepartamento de Recursos Naturais Renováveis, Faculdade de Ciências Agronômicas, Universidade do Estado de São Paulo, Campus de Botucatu, CEP: 18603-970, Botucatu, SP, fax: (014) 8213438, CP.:237
RESUMO
Estudou-se a produção energética na forma de calor da biomassa seca de uma faixa de cerrado sensu stricto da Fazenda Água Limpa, localizada em Brasília-DF. Em dez parcelas, de 20 m x 50 m cada uma, foram identificadas todas as árvores com diâmetro igual ou superior a 5cm, tomado a 30 cm de altura do solo. Foram derrubadas ao acaso, três árvores por espécie em 7 classes diametrais pré-estabelecidas, coletando-se os dados de massa verde do tronco e dos ramos. Em laboratório, a partir de seções transversais, foram separadas madeiras de casca, estabelecendo uma relação mássica. Os teores de umidades da madeira e da casca foram determinados. Desta forma foi possível estimar a massa seca da madeira e da casca, no tronco e nos ramos, por árvore e por área. A comunidade foi caracterizada estruturalmente pelo Índice de Valor de Cobertura. Foram encontrados 47 espécies e 673 indivíduos/ha no povoamento, que se constituiu de 71% de madeira e 29% de casca. A produção média do povoamento foi de 12,38 t/ha e 18,39 kg/árvore. Com base na análise de componentes principais, separou-se um grupo de 17 espécies que foram responsáveis por 83% da produção de biomassa seca por árvore e por 93% da produção de biomassa seca por hectare. Foram determinados os poderes caloríficos superiores e as massas específicas básicas destas espécies. A massa específica variou de 0,38 a 0,78 g/cm3 para a madeira e de 0,41 a 0,67 g/cm3 para a casca. As madeiras apresentaram poderes caloríficos superiores variando de 19.109,09 kJ/kg a 20.883,95 kJ/kg, enquanto para a casca a variação foi de 19.523,50 kJ/kg a 24.019,27 kJ/kg. O poder calorífico médio da madeira foi de 20.025,82 kJ/kg e da casca 20.754,19 kJ/kg. A quantidade média de calor produzida por hectare, foi de 15.207,74 MJ, mas a variação individual entre as espécies foi grande, variando de 290,59 MJ a 58.870,14 MJ. As melhores espécies para a produção de energia na forma de calor foram: Sclerolobium paniculatum (carvoeiro), Dalbergia miscolobium (jacarandá-do-cerrado) e Pterodon pubescens (sucupira branca).
Palavras-chave: Produção de energia, biomassa, madeira, casca, cerrado.
ABSTRACT
The production of energy in the form of heat from dry biomass was studied in a stretch of stricto sensu cerrado scrubland at Água Limpa farm in Brasilia (Federal District, Brazil). In ten plots of land, each measuring 20m x 50m, all the trees with a diameter equal to or larger than 5cm, measured 30cm above ground, were identified. Three trees of each specie were felled at random in seven distinct pre-established diameter categories, data relating to the green mass of the trunk and branches being recorded. In the laboratory, cross-sections were used to separate wood from bark, establishing a mass ratio. The moisture content of the wood and bark were then determined. This made it possible to estimate the dry mass of the wood and bark of the trunk and branches per tree specie and per area. The community was structurally characterised by the Cover Value Index. A total of 47 species and 673 individuals/ha were encountered in the stand consisting of 71% wood and 29% bark. The average production of the stand was 12.38 t/ha and 18.39 kg/tree. After analysing the main components, a group of 17 species were singled out as being responsible for 83% of the production of dry biomass per hectare. The heat combustion and the specific gravity of these species were then determined. The specific gravity varied from 0.38 g/cm3 to 0.78 g/cm3 for wood and 0.41 g/cm3 to 0.67 g/cm3 for bark. The wood presented heat combustion ranging from 19,109.09 kJ/kg to 20,883.95 kJ/kg, whereas the figures for bark ranged from 19,523.50 kJ/kg to 24,019.27 kJ/kg. The average heat combustion of the wood was 20,025.82 kJ/kg compared with 20,754.19 kJ/kg for the bark. The average quantity of heat produced per hectare was 15,207.74 MJ but individual variation among the species was remarkable: 290.59 MJ to 58,870.14 MJ. The best species for the production of energy in the form of heat were: Sclerolobium paniculatum, Dalbergia msicolobium and Pterodon pubescens.
Keywords: Energy production, biomass, wood, bark, "cerrado".
INTRODUÇÃO
No Brasil o consumo de energéticos primários teve nas energias renováveis, no ano de 1998, uma participação de 55,77%. Sendo que 19,71% foi representado pela biomassa, que tem despertado maior interesse como alternativa aos combustíveis fósseis. GRASSI (1994) afirma que: "a produção sustentada de biomassa, que é a fonte mais versátil de energia renovável, apresenta a possibilidade de prover, de forma permanente, grandes quantidades de combustíveis gasosos, líquidos e eletricidade". Ao longo dos últimos 29 anos a lenha, como combustível doméstico, teve papel importante como energético primário no Brasil, como pode ser observado pela Figura 1. Em 1998, dos 19,71% de biomassa, 9,06% foram compostos pela lenha, ou seja, 6,94 x 107 toneladas de madeira foram utilizadas para geração de calor. Do total da lenha consumida, 2,7 x 107 toneladas o foram em forma indireta, principalmente na fabricação de carvão vegetal, e 4,23 x 107 toneladas, em forma direta.
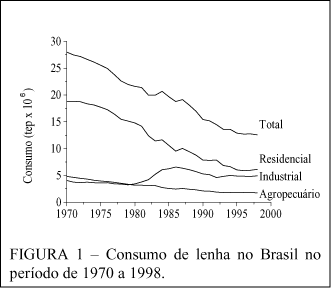
Do total consumido em forma direta, 2,0 x 107 toneladas foram queimadas em residências, para cocção de alimentos (Tabela 1). GOLDEMBERG (1998) afirma que a lenha é, de fato, a fonte de energia dominante nas áreas rurais, e cozinhar é a atividade mais intensiva energéticamente.
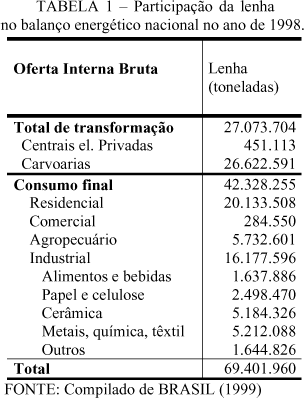
A principal fonte de madeira para produção de energia, no Brasil, tem sido os ecossistemas naturais, com o cerrado e a mata atlântica ocupando os primeiros lugares. O uso mais intensivo da madeira como energético está concentrado nas regiões Sul, Sudeste e Nordeste. A diminuição desses biomas e a pressão conservacionista, associadas à necessidade anual de mais madeira para energia, têm levado a crescente dificuldade para a obtenção desse recurso a partir TABELA 1 – Participação da lenha no balanço energético nacional no ano de 1998.
de florestas nativas. É importante observar que não pesa somente o aspecto quantitativo do uso da madeira como energia, mas também o aspecto econômico, ligado à utilização energética pelas indústrias, e, mais importante ainda, o aspecto social, ou seja, a lenha como energético doméstico (BRITO e DEGLISE, 1991). Nas regiões de uso mais intensivo da madeira como fonte de energia, a situação é de penúria. A população que usa a madeira como fonte de energia não tem condições de obter a quantidade mínima necessária (BRITO, 1990).
Quanto ao aspecto social do consumo da madeira como energético, GOLDEMBERG (1998) sugeriu que o cultivo da lenha em "fazendas energéticas" e seu manejo seriam particularmente significativos para prover uma base de desenvolvimento rural e a geração de emprego em países em desenvolvimento. Enquanto, levando-se em conta a ocupação desordenada do cerrado, OLIVEIRA et al. (1998) propuseram a busca de alternativas para o uso sustentado da vegetação nativa.
O cerrado, um dos cinco maiores tipos de vegetação do Brasil, cobre cerca de 2 milhões de km2 do território (23% do território nacional), com, aproximadamente, 1,5 mlhões de km2 de cerrado sensu lato, localizados no Brasil Central, nos estados de Minas Gerais, Mato Grosso e Goiás (FERRI, 1980). Ocupando uma área densamente povoada, o cerrado é fonte de lenha nativa para a geração de calor e é, talvez, o maior fornecedor de combustível para a cocção de alimentos no meio rural, o que o situa como um dos biomas de grande importância social. Pouco se conhece das espécies do cerrado do ponto de vista energético. Neste contexto uma análise da produção da biomassa seca da comunidade, do poder calorífico e da massa específica da madeira e da casca torna-se importante para um melhor conhecimento do cerrado.
A massa específica é um dos principais índices de qualidade da madeira e, segundo, BRASIL (1972) os métodos que se apóiam na massa específica básica, são os que mais satisfatoriamente medem a quantidade de substância madeira por unidade de volume.
O poder calorífico de um corpo é a quantidade de calor liberada pela combustão de uma unidade de massa desse corpo, e pode ser expresso em calorias por grama ou quilocalorias por quilograma DOAT (1977). A autora definiu, ainda, o poder calorífico superior (PCS) como aquele em que a combustão se efetua a volume constante e no qual a água formada durante a combustão é condensada.
Para BRITO (1986), a variação do poder calorífico superior da madeira está entre 14.561 kJ/kg e 20.930 kJ/kg.
KOLLMANN e CÔTÉ (1968) relataram que o poder calorífico médio para as madeiras situa-se em torno de 18.837kJ/kg a 0% de umidade.
O presente trabalho objetiva estimar a biomassa seca da madeira e da casca das espécies lenhosas de uma faixa de cerrado, indicando as melhores e determinando, para estas, os poderes caloríficos, a massa específica básica e a quantidade de calor disponibilizada.
MATERIAL E MÉTODO
A coleta de dados de campo foi conduzida em uma área de 63,56 ha de vegetação do tipo cerrado sensu stricto, com solo do tipo latossolo vermelho-escuro, segundo SILVA (1999). O clima, segundo classificação de Koppen (NIMER, 1989) é do tipo Aw. A área de estudo localiza-se na Fazenda Água Limpa (FAL), de propriedade da Universidade de Brasília (UnB), Distrito Federal, localizada em altitude de 1.100 metros a 15°56'14''S e 47°46'08''W.
A área foi dividida em parcelas iguais de 20 m x 50 m, dentre as quais sortearam-se, aleatoriamente, 10 parcelas, totalizando uma amostra de 1,57% da área, seguindo procedimento utilizado por SILVA (1990). Todas as árvores com diâmetro igual ou superior a 5 cm medido a 30 cm de altura do solo foram identificadas botânicamente, anotando-se os diâmetros e as alturas totais.
As árvores devidamente identificadas foram distribuídas nas seguintes classes diamétricas, em cm, de (5-9]; (9-13]; (13-17]; (17-21]; (21-25]; (25-29] e (29-33]. Em seguida sortearam-se, ao acaso, três indivíduos por classe diamétrica e por espécie, que foram cortados para a pesagem, no campo, da biomassa aérea e a retirada de amostras da madeira e da casca.
Padronizou-se amostrar a biomassa do tronco, constituída da madeira e da casca das árvores vivas com 5 cm de diâmetro, medido a 30 cm de altura do solo, e da madeira e da casca dos ramos, das mesmas árvores, com diâmetro da base igual ou superior a 3 cm.
Nos indivíduos selecionados e cortados, foram eliminados os ramos finos de diâmetro inferior a 3 cm. O tronco e os ramos maiores remanescentes foram pesados separadamente. Ambas as massas foram obtidas com casca. A seguir, foram recolhidas duas amostras de seções transversais (discos) com aproximadamente 2,50 cm de espessura, ao longo do tronco nas posições correspondentes a 0, 25%, 50%, 75% e 100% da altura do tronco. Foi retirada, também, amostra na base de cada ramo. Em árvores bifurcadas ou com mais de dois ramos principais escolheu-se, ao acaso, um dos ramos para a retirada da amostra. As duas amostras de cada posição, acondicionadas em sacos de polietileno e identificadas, foram encaminhadas ao laboratório para a determinação da umidade, da massa específica básica e do poder calorífico superior respectivamente, para a madeira e para a casca.
No dia da coleta, em laboratório, determinou-se a relação entre a madeira e a casca e os seus teores de umidade para o tronco e os ramos. Para tanto, separou-se a casca da madeira, que foram pesadas para obtenção da massa úmida. Com esses valores, determinou-se a relação mássica entre a madeira e a casca, base úmida, das amostras do tronco e dos ramos.
Esta relação foi utilizada para estimar a massa úmida e seca de madeira e casca por espécie e por área.
Os discos destinados a determinação dos poderes caloríficos superior foram separados em madeira e casca. Cada fração foi misturada numa amostra composta por espécie. As amostras compostas, após serem moídas, foram classificadas em peneiras, obtendo-se a fração retida entre 40 e 60 mesh que foi utilizada para a determinação do poder calorífico superior.
Os teores de umidades foram determinados pelo método da estufa, em base seca, segundo VITAL (1997).
A massa específica básica foi determinada pelo método do máximo teor de umidade, segundo FOELKEL et al. (1971) e VITAL (1984), com amostras retiradas à 25% de altura do tronco em relação ao solo, segundo JESUS et. Al. (1996) e VALE et al. (1999).
Para se verificar o comportamento da espécie no âmbito da estrutura da comunidade, utilizou-se do índice de valor de cobertura (IVC), segundo RODRIGUES (1988), pela soma da abundância relativa e da dominância relativa.
O poder calorífico foi determinado segundo a norma ABNT NBR 8633 (1984) e pelo manual de operações do calorímetro PARR 1201.
A quantidade de calor disponibilizada pela espécie foi obtida pelo produto da massa com o respectivo poder calorífico para cada espécie.
Uma análise de componentes principais foi feita para separar espécies de maior produção de biomassa seca, segundo CRUZ e REGAZZI (1994).
RESULTADOS E DISCUSSÃO
Foram encontrados 673 indivíduos por hectare distribuídos entre 47 espécies e 24 famílias. Em estudos conduzidos em área similar de cerrado, SILVA (1990) encontrou 35 espécies e 1.233 árvores/ha e cita OLIVEIRA et. al. (1982), que encontraram 35 espécies e 567 árvores/ha. Em outras áreas de cerrado, GOODLAND (1971) levantou 43 espécies e 2.253 árvores/ha; RIZZINI (1979), 29 e 847; MEDEIROS (1983), 38 e 947; RIBEIRO e HARIDASAN (1984), 50 e 550; RIBEIRO et al. (1985), 51 e 730. O povoamento apresentou um diâmetro médio de 11,39 cm a 30 cm de altura em relação ao solo, variando de 5,00 cm a 32,30 cm., com a altura total média de 4,10 m, variação de 1,00 m a 14,00 m. Os valores médios de diâmetro e altura foram ponderados pelo número de indivíduos por classe diamétrica.
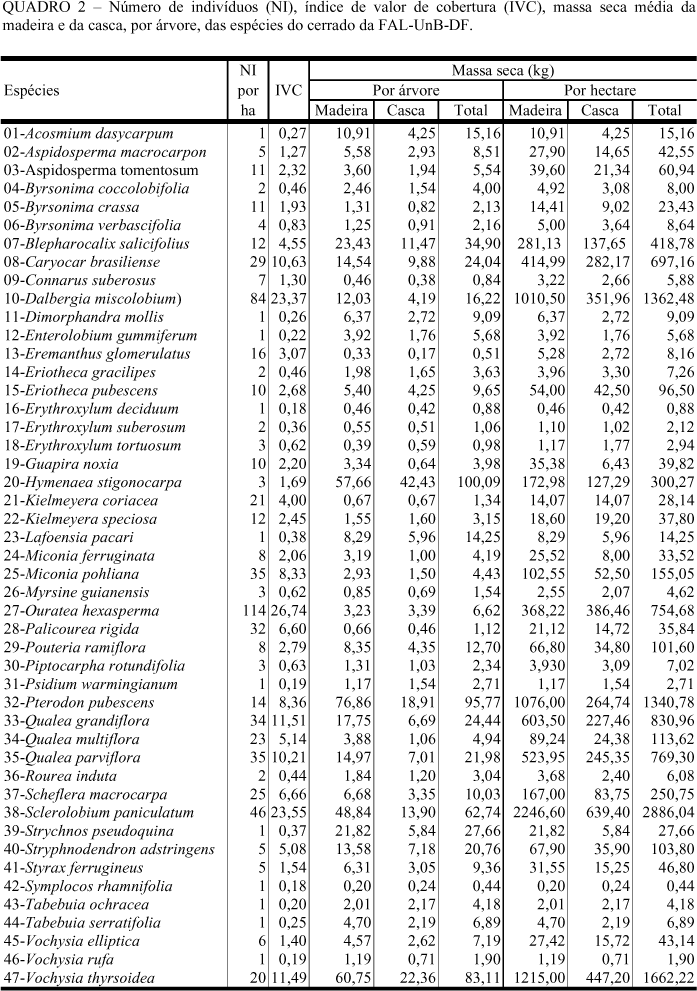
A biomassa originada pelo cerrado da Fazenda Água Limpa, da UnB - DF, estimada na forma de massa seca por indivíduo e massa seca por hectare, para cada espécie, está na Tabela 2.
A produção média, ponderada pelo número de indivíduos por classe diamétrica, foi de 18,39 kg/árvore e de 12,38 toneladas/ha, produção considerada baixa se comparada à de outros ecossistemas florestais.
OLIVEIRA (1993) citando vários trabalhos apresenta para floresta subtropical, produções de 213,50 t/ha a 513,00 t/ha; para floresta tropical úmida, produções de 204,50 t/ha a 240,90 t/ha; para floresta alagada, de 172,60 t/ha; para campina amazônica, de 77,90 t/ha; para floresta de galeria, de 155,90 t/ha a 199,00 t/ha, para fitomassa aérea seca de cerrado de 34,8 a 46,9 t/ha. Dados compilados de SILVA (1990), para massa seca de tronco e ramos de uma área de cerrado do Distrito Federal, mostraram uma produção de 17,29 t/ha.
Na escolha de árvores para produção de biomassa energética deve-se optar por espécies que tenham características energéticas favoráveis, como alto poder calorífico e boa massa específica básica, entre outras. Aliado a estas características, a espécie deve ser boa produtora de biomassa.
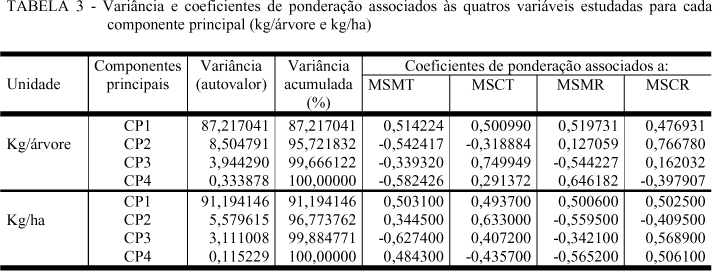
Desta forma separaram-se as espécies em função de suas massas secas individuais e por hectare, utilizando-se de análise de componentes principais, tendo como variáveis independentes a massa seca de madeira do tronco (MSMT), a massa seca de casca de tronco (MSCT), a massa seca de madeira de ramos (MSMR) e a massa seca de casca dos ramos (MSCR), cujos resultados são apresentados na Tabela 3.
CRUZ e REGAZZI (1994), afirmam que os primeiros componentes principais têm sido utilizados quando envolvem, pelo menos, 80% da variação total. Pode-se observar pela Tabela 3, que os primeiros componentes principais explicaram juntos, 95,72% e 96,77%, respectivamente para produção individual e por hectare, da variação total das espécies em relação às massas secas.
Considerando a produção individual e utilizando os dois primeiros componentes principais para identificar os grupos mais homogêneos de espécies pode-se, distribuindo os valores de seus escores num eixo cartesiano (xy), observar a dispersão dos pontos e a formação de grupos. Desta forma, destacou-se (Figura 2) um primeiro grupo formado por 5 espécies: Blepharocalix salicifolius, Pterodon pubescens, Sclerolobium paniculatum, Vochysia thyrsoidea e Hymenaea stigonocarpa, cujos números de referência são 7, 32, 38, 47 e 20.
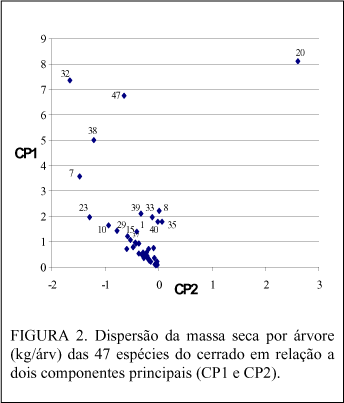
O segundo grupo foi formado por 11 espécies: Lafoensia pacari (23), Dalbergia miscolobium (10), Pouteria ramiflora (29), Eriotheca pubescens (15), Acosmium dasycarpum (1), Strychnus pseudoquina (39), Caryocar brasiliensis (8), Qualea grandiflora (33), Stryphonodendrum adstringens (40), Qualea parviflora (35) e Scheflera macrocarpa (37). As 31 espécies restantes formaram o terceiro grupo.
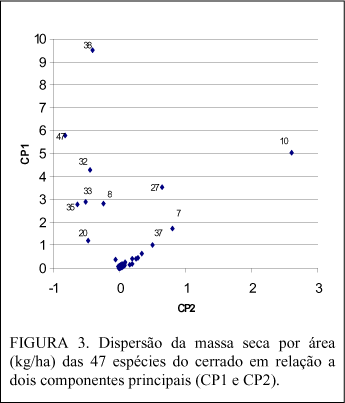
Considerando-se o mesmo critério anterior, para a produção por hectare, o destaque (Figura 3), num primeiro grupo formado por 5 espécies, foi para Sclerolobium paniculatum (38), Dalbergia miscolobium (10), Vochysia thyrsoidea (47), Pterodon pubescens (32) e Ouratea hexasperma (27). O segundo grupo foi formado por seis espécies: Qualea grandiflora (33), Blepharocalix salicifolius (8), ), Qualea parviflora (35), Caryocar brasiliensis (8), Scheflera macrocarpa (37) e Hymenaea stigonocarpa (20). As 36 espécies restantes formaram o terceiro grupo.
Em ambos os casos, o componente principal 1 (CP1) explicou sozinho mais de 85% da variação.
Quando se considerou a produção individual, o componente principal 1 (CP1) explicou 87,217041% da variação, tendo a massa seca da madeira de ramo (MSMR) a variável de maior peso (0,519731), seguida da massa seca da madeira de tronco (MSMT), cujo valor foi de 0,514224. O componente principal 2 (CP2) explicou apenas 8,504797% da variação e teve a massa seca da casca de ramos (MSCR) com maior peso (0,766780).
Quando se considerou a produção de massa seca por hectare (Figura 3), a massa seca da madeira de tronco e de ramo juntamente com a massa seca da casca de ramo tiveram praticamente os mesmos pesos para a componente principal 1 (CP1), que, por sua vez, explicou 91,194146% da variação, enquanto a componente principal 2 (CP2) explicou apenas 5,579615% da variação tendo a massa seca da casca de tronco a variável de maior peso.
Observa-se, pela Figura 2 e 3, que dezessete espécies destacaram-se quanto à produção de biomassa sendo que algumas se apresentaram com boa produção tanto individual quanto por área.
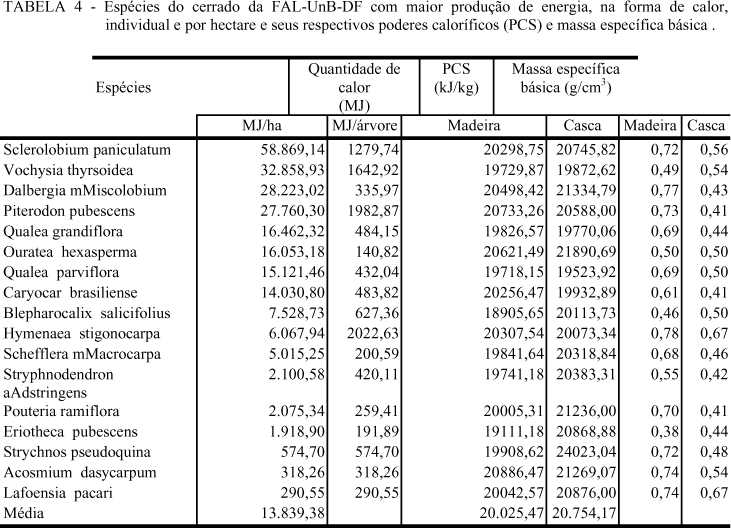
A Tabela 4 apresenta essas espécies com as respectivas características físicas e energéticas de sua madeira e de sua casca.
Acosmium dasycarpum foi a espécie que apresentou o maior poder calorífico para a madeira e um dos maiores para casca. A espécie teve também alta massa específica básica. É, entretanto, como a Lafoensia pacari, espécie de pequeno porte que produz baixa quantidade de energia, em função da pequena produção de biomassa seca. Strychnos pseudoquina também pode ser incluída nesse grupo, apesar da produção energética maior (Tabela 4). Na observação dos índices de valor de cobertura (Tabela 2), verificou-se que têm pequena participação na estrutura da comunidade, não sendo, portanto, importantes na produção energética da comunidade.
Eriotheca pubescens e Stryphnodendron adstringens tiveram baixo índice de valor de cobertura (Tabela 2) possuindo menores valores de poder calorífico superior da madeira e de massa específica básica (Tabela 4), contribuindo muito pouco para a produção energética.
Blepharocalix salicifolius e Vochysia thyrsoidea apresentaram madeiras com baixas massas específicas básicas e baixos poderes caloríficos e, embora com boa produção de biomassa seca, produziram pouca energia por unidade de massa e pouca energia por unidade de volume. Os valores de IVC foram de 4,55 e 11,49, respectivemente, para Blepharocalix salicifolius e Vochysia thyrsoidea (Tabela 2).
Caryocar brasiliense apresentou madeira com massa específica média e alto poder calorífico e na comunidade, 10,63 de valor de IVC (Tabela 2). Esteve entre as espécies consideradas medianas na produção total de energia.
Qualea grandiflora, Qualea parviflora e Scheflera macrocarpa apresentaram massas específicas médias e, respectivamente, 11,51; 10,21 e 6,66 de IVC, (Tabela 2). Essas espécies tiveram, poderes caloríficos e produções energéticas iguais ou abaixo da média. São árvores de pequeno porte, e não devem receber prioridade na escolha de espécies para produção de energia.
Com massas específicas e poderes caloríficos superiores acima da média encontrou-se Hymenaea stigonocarpa e Pouteria ramiflora, sendo a primeira uma das maiores produtoras individuais de biomassa seca (Tabela 4). Do ponto de vista da comunidade, a espécie H. stigonocarpa teve um valor muito baixo de IVC (1,69). P. ramiflora gerou um IVC de 2,79 (Quadro 2). Foram espécies com produção total de energia muito abaixo da média da comunidade.
Ouratea hexasperma foi a espécie com o maior índice de valor de cobertura (26,74), em função do maior número de indivíduos/ha da comunidade (Tabela 2). A produção total de calor situou-se na média (Tabela 4). As árvores são de pequeno porte.
Sclerolobium paniculatum, Dalbergia miscolobium e Pterodon pubescens foram as melhores espécies tanto ao nível individual, se analisadas as características da madeira para produção de energia, quanto no da comunidade. Possuem, as duas primeiras, altos índices de valor de cobertura (Tabela 2).
Sclerolobium paniculatum, além da alta massa específica e alto poder calorífico, mostrou boa produção de biomassa seca. Foi, dentre as espécies da comunidade, a que apresentou o segundo maior IVC (Tabela 2). Foi, também, a espécie que produziu quase o dobro de energia por hectare se comparada com a espécie imediatamente abaixo (Tabela 4).
Dalbergia miscolobium, com IVC semelhante a Sclerolobium paniculatum, apresentou o dobro do número de indivíduos em relação a ele. (Tabela 2).
Pterodon pubescens, com IVC menor (8,36), com alto poder calorífico e a segunda maior produção individual de massa seca teve uma produção energética semelhante à Dalbergia miscolobium.(Tabela 4).
Pode-se distinguir no cerrado três categorias de espécies. Na primeira estariam as espécies com características individuais inferiores para a produção de energia mas com alta produção de biomassa. Na segunda categoria estariam as espécies que não têm participação efetiva na estrutura da comunidade, ocorrendo em menor número por hectare. São espécies que não teriam destaque numa possível exploração sustentada. Ao terceiro grupo pertencem as espécies que tanto nas características individuais quanto na produção por área possuem as melhores características para utilização num programa sustentado de produção de combustível doméstico.
CONCLUSÃO
Os 673 indivíduos encontrados por hectare, pertencentes a 47 espécies, mostraram grande diversidade de respostas aos parâmetros escolhidos para indicação de produção de energia, na forma de calor.
A produção média de biomassa seca total para a área foi de 12,38 t/ha, com variações individuais de 0,44 kg/ha (Symplocos rhamnifolia com 1 indivíduo por ha) a 2886,04 kg/ha (Sclerolobium paniculatum com 46 indivíduos por ha). Em função da estrutura da comunidade, houve espécies com boa produção individual mas com valores muito baixos por hectare. A produção média por árvore foi de 18,39 kg.
A massa específica básica média da madeira variou de 0,20 g/cm3 a 0,78 g/cm3, e da casca de 0,17 g/cm3 a 0,67 g/cm3. A espécie de maior massa específica básica para a madeira e para a casca foi Hymenaea stigonocarpa. O poder calorífico superior médio da madeira foi de 9.942,10 kJ/kg, e o da casca, de 20.264,42 kJ/kg.
A quantidade de calor média por hectare produzida, entre as dezessete espécies que se destacaram, foi de 15.209,16 MJ, mas a variação individual entre as espécies foi grande. Sclerolobium paniculatum originou 58.869,14 MJ, enquanto Lafoensia pacari, apenas 290,55 MJ.
A variabilidade na comunidade mostrou espécies com grande produção energética em função da alta produção de biomassa porém com maderia sem as características desejáveis para a geração de calor. Dentre elas, está Vochysia thyrsoidea, que produz grande quantidade de energia por hectare (32.858,92 MJ). Têm-se também espécies com boas características da madeira mas que não se destacam na comunidade para produção energética, em função de baixa produção de biomassa. Acosmium dasycarpum, com apenas um indivíduo por hectare, está entre essas espécies. Houve espécies com boas características da madeira e com alta produção de biomassa seca, que originaram grande quantidade de energia (Sclerolobium paniculatum).
Podem-se distinguir três categorias de espécies na comunidade quanto à geração de energia. Na primeira, estariam as espécies em que as baixas características da madeira para produção de energia deveriam ser compensadas com volume maior de material para ser queimado, o que implica maior consumo de madeira para gerar a mesma quantidade de calor. Na segunda categoria se colocariam as espécies que não têm grande participação na estrutura da comunidade, ocorrem em menor número por hectare, mas que não podem ser esquecidas em estudo que vise à produção de energia, pois apresentam boas qualidades individuais da madeira. Ao terceiro e último grupo pertenceriam as espécies com excelentes características, tanto na produção de biomassa seca quanto na qualidades de suas madeiras, destacando-se entre as maiores produtoras de energia. São elas: Sclerolobium paniculatum, Dalbergia miscolobium e Pterodon pubescens.
REFERENCIAS
[2]ABNT – Associação Brasileira de Normas Técnicas; NBR 8633; 13p; 1984.
[4]BRASIL, Ministério de Minas e Energia; Balanço Energético Nacional; Brasília; 153p.; 1999.
[5]BRASIL, M.A.M; Escola Superior de Agricultura Luiz de Queiroz – USP; Variação da densidade básica da madeira de Eucalyptus propinqua Deane ex Maiden em função do local e do espaçamento; Piracicaba; 75p.; 1972. (Dissertação de Mestrado).
[8]BRITO, J.O. e DEGLISE, X.; ESALQ-USP; States and potencial of using wood for energy in Brazil; In: 10° World Forest Cogress; Actes Proceedings Actas 6; Revue Forestière Française; n.6; p.175-79; Paris; 1991.
[9]BRITO, J.O.; ESALQ-USP; Forest Resources Contribution to energy splies in Brazil; IPEF Internacional; n.1; p.23-25; Piracicaba; 1990.
[10]BRITO, J.O.; ESALQ-USP; Madeira para energia: a verdadeira realidade do uso de recursos florestais; In: 5° Congresso Florestal Brasileiro; Anais; Sociedade Brasileira de Silvicultura; p.188-193; Recife; 1986.
CRUZ, C. D., REGAZI, A.; UFV; Moldelos biométricos aplicados ao melhoramento genético; Imprensa Universitária; 390p.; Viçosa; 1994.
[13]DOAT, J.; Le puovoir calorifique des bois tropicaux. Revue Bois et Forêts des Tropiques; n.172,; p.33-48; França; 1977.
FERRI, M.G.; USP; Vegetação Brasileira. Ed. Itatiaia, Belo Horizonte; Ed. da Universidade de São Paulo; São Paulo; 157p.; 1980.
[14]FOELKEL, C.E.B., BRASIL, M.A.M. e BARRICHELO, L.E.G; Métodos para determinação da densidade básica de cavacos para coníferas e folhosas; IPEF; n.2/3; p.65-74; Piracicaba; 1971.
[15]GOLDEMBERG, J; USP; Energia, Meio Ambiente & Desenvolvimento; Editora da Universidade de São Paulo; 125p.; São Paulo; 1998.
GOODLAND, R. Oligotrofismo e alumínio no cerrado; In: 3° Simpósio sobre o cerrado; Anais; Editora Universidade de São Paulo e Edgard Blucher; p.44-60; São Paulo; 1971.
[16]GRASSI, G., PALZ, W.; O futuro da biomassa na União Européia; Álcool. & Açúcar; n.76; p.28-34; 1994.
[20]JESUS, R.M., VITAL, B.R.; UFV; Comparação entre densidades de matrizes e de clones de Eucalyptus grandis; Revista Árvore; v.10; n.1; p.83-90; Viçosa; 1996.
[21]KOLLMANN, F. e CÔTÉ Jr., W.A. Principles of wood science and technology. I - solid wood; Springer Verlag; 592p.; New York; 1968.
MEDEIROS, R.A; Comparação do estado nutricional de algumas espécies acumuladoras e não acumuladoras de alumínio, nativas do cerrado; Departamento de Biologia Vegetal; Universidade de Brasília; 91p.; Brasília; 1983.; (Dissertação de Mestrado)
NIMER, E.; Climatologia do Brasil. Departamento de Recursos Naturais e Estudos Ambientais; IBGE; Rio de Janeiro; 1989.
OLIVEIRA, A.D., LEITE, A.P., BOTELHO, S.A., SCOLFORO, J.R.S.; ESAL; Avaliação econômica da vegetação de cerrado submetido a diferentes regimes de manejo e de povoamentos de eucalipto plantado em monocultivo. CERNE,; v.4; n.1; p.34-56; Lavras; 1998.
RIBEIRO, J.F., HARIDASAN, M.; EMBRAPA -UnB; Comparação fitossociológica de um cerrado denso e um cerradão em solos distróficos no Distrito Federal. In.: 35° Congresso nacional de botânica; Anais; Sociedade Botânica do Brasil; p. 342-353; Manaus; 1984.
RIBEIRO, J.F.; SILVA, J.C.S., BATMANIAN, G.J.; EMBRAPA; Fitossociologia de tipos fisionômicos de um cerrado em Planaltina-DF; Revista Brasileira de Botânica; n.8; p.131-142; 1985.
RIZZINI, C.T. ; Tratado de fitogeografia do Brasil – Aspectos sociológicos e florísticos; Editora Universidade de São Paulo; São Paulo; 1979.
[24]RODRIGUES, R.R. ESALQ-USP; Métodos fitossociológicos mais usados; Casa da Agricultura; ano 10; n°1; 1988.
SILVA, M.A.; Mudanças na composição florística e estrutura de um cerrado sensu stricto em um período de 12 anos (1985-1997), na Fazenda Água Limpa (FAL) – Distrito Federal; Faculdade de Tecnologia; Departamento de Engenharia Florestal; Universidade de Brasília; 62p; Brasília; 1999; Dissertação de Mestrado em Ciências Florestais)
[25]SILVA, F.C. EFL-UnB; Compartilhamento de nutrientes em diferentes componentes da biomassa aérea em espécies arbóreas de um cerrado; Instituto de Ciências Biológicas; Departamento de Ecologia; Universidade de Brasília; 80p.; Brasília; 1990; (Dissertação de Mestrado em Ecologia).
[27]VALE, A.T., BRASIL, M.A.M., MARTINS, I.S. EFL-UnB; Variação axial da densidade básica da madeira de acacia mangium willd aos sete anos de idade; Revista Ciência Florestal; v.9; n. 2; 1999.
[31]VITAL, B.R.; DEF-UFV; Métodos de determinação da densidade da madeira; Boletim Técnico; n.2,; 21p; Viçosa; 1984.
[32]VITAL, B.R. DEF-UFV; Métodos para determinção do teor de umidade da madeira; Boletim Técnico; n. 13; 33 p.; Viçosa; 1997.
Endereço para correspondência
Ailton Teixeira do Vale
e-mail: atvale@unb.br
{kind=link}
{kind=link}
{kind=link}